诺奖得主|斯万特·帕博:基因流动
按:本文节选自《尼安德特人》(浙江教育出版社出版,后浪出品,2018年12月)一书。作者斯万特·帕博(Svante Pääbo),2022年诺贝尔生理学或医学奖获得者,1955年出生于瑞典斯德哥尔摩,1986年在乌普萨拉大学取得博士学位,后在瑞士苏黎世大学和美国加州大学伯克利分校从事博士后研究。1990年,他成为德国慕尼黑大学教授,并于1999年在德国莱比锡创立了马克斯普朗克进化人类学研究所,至今仍活跃在该研究所,同时他还是美国、瑞典等国的国家科学院院士。其父苏恩·伯格斯特龙是1982年的诺贝尔生理学或医学奖得主。

2009 年 5 月,我们开始对 5 个现代人的基因组进行测序。这些原始 DNA 没有受到像尼安德特人样品那样的细菌污染和化学损伤,所以我们从每一个样品中得到的 DNA 序列数量约为尼安德特人的 5 倍。一两年前,我们还无法想象在莱比锡测序这些基因组。不过454 和宜曼达这种公司的测序技术一推向市场,我们这样的小型研究团队就能够在短短几周内完成几个人的全基因组测序任务。
艾德使用他在杜布罗夫尼克描述过的方法,估计这 5 个现代人的基因组与人类参考基因组在多久之前拥有共同的祖先。他发现,欧洲人、巴布亚人和中国人与人类参考基因组在 50 多万年前有着共同的祖先。如果加入来自南非的桑河人(San),那么谱系分开的时间就在将近 70 万年前。桑河人(及相关的种群)和其他非洲人、世界上其他地方的人分开的时间最久。因此,尼安德特人和现代人基因组的共同祖先约在 83 万年前,只比桑河人与人类参考基因组的共同祖先早了 13 万年。所以尼安德特人确实与我们不同,但差别不是很大。
我们必须谨慎对待这样的计算,因为它们只提供了一个共同祖先的年代值,而这个年代值似乎对整个基因组都是一样的。但是,基因组并非一个整体的遗传单位。这意味着每个个体基因组的每个部分都有自己的历史,因此每个部分和其他基因组也有不同的共同祖先。这是因为每个人都携带每条染色体的两份拷贝,其中一份拷贝会独立地传递给孩子。因此,每一条染色体都有独特的历史模式,或自身系谱。此外,在卵细胞和精子形成之时,每一对染色体都以错综复杂的分子舞蹈相互交换片段,这种方式又称为重组。因此,不仅群体中每条染色体都有自己的系谱,每条染色体的每个片段也有。因此,艾德用人类参考基因组计算出来的共同祖先的年代,无论是与尼安德特人的共同祖先在 83 万年前,还是与桑河人的共同祖先在 70 万年前,这些数字都只代表了基因组所有部分的平均值。
事实上,当我们将两个现代人的 DNA 区域进行相互比较时,很容易发现有些区域的共同祖先在几万年前,而有些区域的共同祖先在 150 万年前。当我们比较现代人基因组与尼安德特人基因组时,也会发现这种结果。如果有人能在我的一条染色体上散步,并将它与尼安德特人的染色体以及正在阅读这本生命基因之书的读者的染色体进行比较,那么染色体漫步者会发现,有时候我与尼安德特人更相似,有时候读者与尼安德特人更类似,有时读者和我更相似。艾德的平均值只意味着,在比对读者、我与尼安德特人的基因组时,读者和我之间有着更多更相似的基因组区域。
83 万年只是现代人类 DNA 序列与尼安德特人化石所携带的 DNA 序列拥有共同起源的平均时间,意识到这点十分重要。这些 DNA 序列最早存在于一个种群身上,后来该种群的后代成了尼安德特人及现代人类的祖先。但这并不是现代人类和尼安德特人彼此分开的那个时间点,分离肯定是后来才发生的。因为我们追溯了现今人类的 DNA 序列历史和尼安德特人的 DNA 序列历史,发现这两个世系首先会进入现代人类和尼安德特人最后的祖先群(之后分离形成的两个种群就源于此),然后再进入这个祖先群当时的变异之中。所以 83 万年是一个复合时间,它包括尼安德特人和现代人类成为不同种群的时间,还包括它们共同祖先群遗传变异的时间。
对我们而言,祖先的种群仍然很神秘,虽然我们认为它曾居住在非洲,后来其后代有一些离开那里,成为尼安德特人的祖先,而那些留下来的人则成了当今人类的祖先。用DNA 序列来估计这两个种群的分开时间很棘手,比用 DNA 序列来估计它们共同祖先的时间要难得多。例如,如果尼安德特人和现代人类的祖先群间存在很多变异,若我们在祖先群中积累的 DNA 序列差异多于尼安德特人和现代人类分道扬镳之后的差异,那就说明这两个群体的分离时间相对新近。我们可以估计基因组不同片段的演化时间,以此推测差异性,并粗略估计出祖先种群的变异水平。为估计种群的分开时间,我们还需要知道世代时间,也就是每个个体产下后代的平均年龄。但对于这个时间,我们显然一无所知。我们竭尽所能把这些不确定性都纳入考虑,得出的结论是,这两个种群大约在 44 万至 27 万年前的某个时候就分开了。当然,我们可能还是低估了许多不确定性。不过,当今人类的祖先与尼安德特人的祖先至少在 30 万年前便已分道扬镳。
估算尼安德特人与现代人类的差异之后,我们要回到原来的问题:当现代人类的祖先离开非洲,并在欧洲遇到失散已久的尼安德特人“表兄弟”之后,他们之间发生了什么。为了了解那些现代人类和尼安德特人之间是否存在基因交换,艾德将那 5 个现代人的基因组与黑猩猩的基因组进行比对,并且戴维和尼克也重复了他们自己的分析。我相信,这次结果是可靠的,我之前暗自怀疑的尼安德特人与欧洲人、中国人之间的特别相似之处应该会消失。
7 月 28 日,我收到了来自戴维和尼克的两封长邮件。这很好地见证了戴维对科学的热情,因为他的妻子尤金妮娅(Eugenie)刚刚于 7 月 14 日生下他们的第一个孩子,他仍马不停蹄地着手分析工作。尼克对 5 个现代人类基因组进行了 10 种可能的两两比较。在每种情况下,他均确定了两个个体之间同一染色体存在差异的 SNP。他在任一对基因组之间均发现了大约 20 万个这样的差异。这么多 SNP 足够让我们准确地确定尼安德特人更接近于哪个人。
尼克发现,尼安德特人与桑河人的匹配值为 49.9%,与巴布亚人样本的匹配值为50.1%。这都在预料之中,毕竟尼安德特人从未出现在非洲,因此相较于其他人,尼安德特人与非洲人的关系不会更密切。他使用桑河人与法国人的 SNP 时发现,尼安德特人与法国人的匹配值为 52.4%。我们现在拥有大量数据,所以这些值的不确定性只有 0.4%。因此很明显,与桑河人的基因组相比,法国人与尼安德特人更为相似。他采用法国人和约鲁巴人的 SNP 进行比较时发现,相应的匹配值为 52.5%。采用中国人与桑河人及中国人和约鲁巴人的 SNP 时,相应的匹配值分别为 52.6% 和 52.7%;采用巴布亚人与那两个非洲人的 SNP时,匹配值分别是 51.9% 和 52.1%。采用法国人、中国人和巴布亚人相互间的 SNP 进行分析时,得到的匹配值在 49.8%~50.6% 间变化。因此,在所有不涉及非洲人的比较中,匹配值均为 50% 左右。但每当把非洲人和非洲以外的人进行比较时,尼安德特人与非洲以外的人匹配的 SNP,比尼安德特人与非洲人匹配的 SNP 多 2%。所以无论他们曾住在哪里,尼安德特人对非洲以外的人,确实有小而明显的遗传贡献。
我读了一遍那两封电子邮件,然后又读了一遍。这一遍我读得很仔细,试图找出任何分析上的问题,但一无所获。我靠在办公椅上,茫然看着凌乱的书桌,上面堆满了过去几年的论文和笔记,一层摞一层。电脑屏幕上显示着戴维和尼克得到的结果,直勾勾地盯着我。这次不是由于某种技术错误,而是尼安德特人真的为现代人类贡献了 DNA。这真的超级酷。这是我过去 25 年一直梦想得到的答案。关于人类起源的问题已经争议了几十年,我们现在有确凿的证据来回答这个问题,而且答案出人意料。并非所有现今人类的基因组信息均可追溯到非洲新近的祖先,这与严谨的“走出非洲”假说相矛盾,而我的导师艾伦·威尔逊曾是“走出非洲”的主要构建者之一。这些证据也与我自己长期以来坚信的理论背道而驰。尼安德特人没有完全灭绝,他们的 DNA 仍存在于今天的人类中。
我茫然地盯着桌子,意识到我们结果的出乎意料并不只是因为与“走出非洲”的假设相矛盾,还因为它也不支持“多地区起源”假说。与“多地区起源”假说的预测相反,我们不只在欧洲人中发现了尼安德特人的遗传贡献,在中国人和巴布亚新几内亚人中也看到了相应的证据。这怎么可能呢?我开始心不在焉地整理桌子。起初动作很慢,但越来越振奋。我丢掉多年积累的项目材料,桌上的几摞书之间灰尘飞扬。我需要开启新的篇章。我需要一张干净的桌子。
整理有时能帮助思考。清理完毕后,我在地图上画出箭头,直观地标示出现代人类走出非洲,并和尼安德特人在欧洲相遇的可能路径。我可以想象他们与尼安德特人生下小孩,接着这些孩子融入现代人类,但我想不明白尼安德特人的 DNA 是如何到达东亚的。这很可能是现代人类随后迁移,然后将尼安德特人的 DNA 带入中国。如果真是这样,那么相较于欧洲人和尼安德特人的相似程度,中国人和尼安德特人之间的相似度应该更低。但事实并非如此。然后我意识到:根据我想象的箭头,现代人类走出非洲后经过了中东!这应该是现代人类与尼安德特人相遇的首个地方。如果这些现代人类与尼安德特人杂交,然后成了今天所有非洲以外的人的祖先,那么每个非洲以外的人所携带的尼安德特人 DNA量应该都相同(见下图)。这种情况应该有可能存在。但经验告诉我,有时我的直觉正好是错误的。幸运的是,我知道,如果我错了的话,像尼克、戴维和蒙蒂这样用数学思维验证理论的人会纠正我。
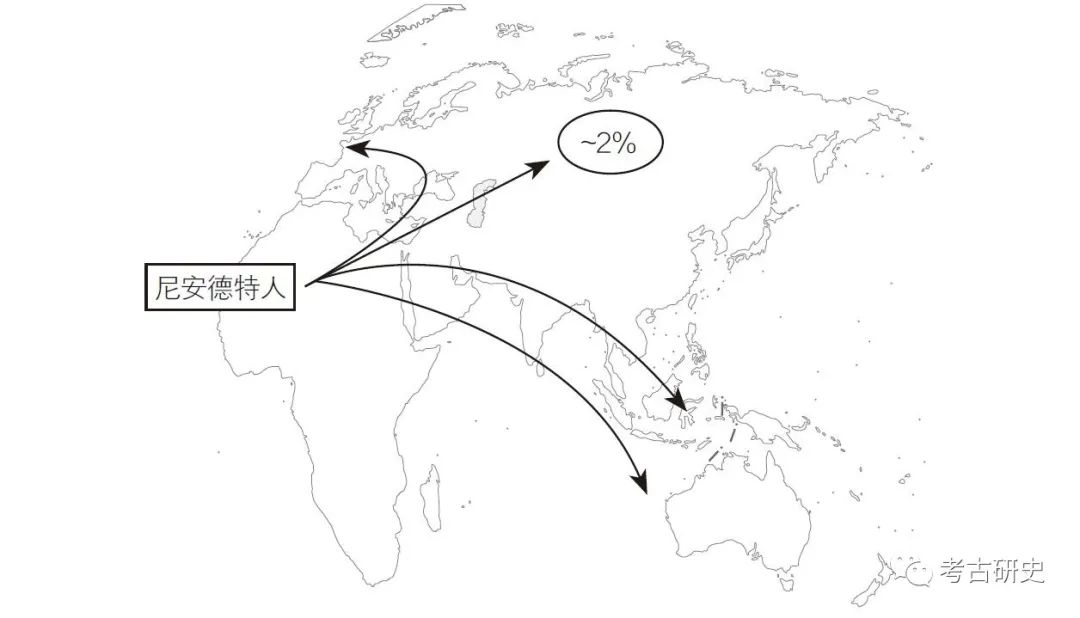
尼安德特人的杂交设想图。如果离开非洲的早期现代人类和尼安德特人杂交后,继续在世界上非洲以外的各地繁衍,那么他们将把尼安德特人的 DNA 携带到尼安德特人从未居住过的区域。例如,中国人的 DNA 中有约 2% 来自尼安德特人。
我们在每周五的会议上以及紧张的联盟电话会议中讨论了戴维和尼克的研究结果。现在我们中的一些人相信,尼安德特人与现代人类有杂交,但也有人仍然不愿意相信,并且他们试图去指出戴维和尼克的分析可能错在何处。我意识到,如果让联盟内的每个人相信这些结果都那么困难的话,要让全世界人都信服就更难了,尤其是许多古生物学家并未在化石记录中发现过现代人类与尼安德特人杂交的证据。这其中包括不少古生物领域内很受尊敬的人,如伦敦自然历史博物馆的克里斯·斯特林格和加州斯坦福大学的理查德·克莱因(Richard Klein)。虽然我认为这些古生物学家在解释化石记录时非常谨慎,但是仍可能在研究过程中受先前遗传结果的影响。包括我们在内的许多研究团队已经表明,现今人类所有的遗传变异,均是来自非洲的新近遗传变异。我们在 1997 年的那篇论文中表明,尼安德特人没有给现今人类贡献任何线粒体 DNA。这篇论文的影响同样深远。虽然密歇根大学的米尔福德·沃尔波夫和华盛顿大学-圣路易斯大学的埃里克·特林考斯等古生物学家在化石中发现了杂交的证据,以及一些遗传学家也曾试图指出现代人类的基因变异可能来自尼安德特人,但这些论点不足以动摇学界的普遍认识。至少他们没有给我造成强烈的冲击。以前从不需要借助尼安德特人的遗传贡献来解释当今世界人类的形态或遗传变异模式。但是现在不同以前,我们可以直接透过尼安德特人基因组来研究。我们看到了遗传贡献,虽然很少。
不过,我觉得还需要得到更多证据才能让世界信服我们的结果。科学并非只是客观、不偏不倚地寻找到科学家无法想象的、颠扑不破的真理。事实上,科学研究是一项社会工作,其中位高权重的人和具有影响力的学者所主张的教条,经常决定了科学“常识”。要打破这些常识,我们必须开展更多关于尼安德特人基因组的分析,不同于戴维和尼克计算SNP 等位基因的方法。如果更多的独立证据也表明,从尼安德特人到现代人类确实存在基因流动,那么说服全世界大多数人就会变得更加容易。因此,寻找其他的可行分析成了我们每周电话会议新的恒定主题。
有点出乎意料的是,可行的建议来自我们联盟外部。在 2009 年 5 月的冷泉港会议上,戴维遇到了拉斯穆斯·尼尔森(Rasmus Nielsen)。拉斯穆斯是一名丹麦的群体遗传学家,曾于 1998 年与蒙蒂·斯拉特金一起完成博士论文。他现在是加州大学伯克利分校的群体遗传学教授。拉斯穆斯告诉戴维,他和他的博士后翟巍巍按区域搜寻了当今人类的基因组,发现非洲之外的变异比非洲之内的更大。虽然这很有可能,但这种模式通常是我们想象不到的,因为从较大种群中分离出来的小分支,一般只包含祖先种群中的一部分变异。如果发现了这样的区域,那么有许多种解释,但有一种可能让我们非常感兴趣。与现代人类分开之后,尼安德特人在非洲之外独立生活了数十万年,一定积累了不同于现代人类的遗传变异。如果他们随后贡献了基因组片段给非洲以外的人,那么采用拉斯穆斯的方法很可能找到这些基因组区域,因为这些区域的遗传模式,正好是非洲之外的变异比非洲之内的更多。使用手头的尼安德特人基因组,我们现在可以检查这些区域是否有一部分基因来自尼安德特人,因为在拉斯穆斯研究的区域中,非洲以外的 DNA 序列更接近于尼安德特人。2009 年 6 月,我邀请拉斯穆斯和巍巍加入尼安德特人基因组分析联盟。
拉斯穆斯专注研究那些很早便与非洲人分开的不同寻常的欧洲区域。他一共发现了 17个这样的区域。艾德把 15 个位于这些区域的尼安德特人 DNA 序列寄给拉斯穆斯。后者于7 月回了信,结果令人吃惊。在这 15 个区域中,有 13 个区域含有存在于欧洲人,但不存在于非洲人中的尼安德特人变异。后来,拉斯穆斯进一步细化了分析结果,并主要集中在DNA 序列超过 10 万个核苷酸的 12 个区域。他发现在其中的 10 个区域中,尼安德特人携带的突变仍能在如今的欧洲人身上找到。这确实是一个惊人的结果!除了解释为基因从尼安德特人传到非洲以外的人,我想象不出其他可能。虽然这只是科学家常挂在嘴边的定性结果,无法凭此计算出尼安德特人对欧洲人或亚洲人的 DNA 贡献,不过它生动地证明了遗传贡献的存在。这种方法虽然与定量分析不同,但它独立验证了戴维和尼克的方法,而且得到了同样的结论。
我们继续思考其他检测基因流的方法。戴维一如既往地想出了绝妙的点子。他的想法很简单:现今人类基因组的一些区域与尼安德特人的相似,因为突变很少,可能是由于突变率低,或由于突变会导致个体死亡。如果我的某个基因组区域也因为同样的原因而与尼安德特人相似,那么我的这个区域也可能与现今其他人类的相近,因为它很少改变。但如果我的基因组区域与尼安德特人相似是因为我的祖先从尼安德特人那里继承了该区域,那么我的这个基因组区域就没有理由和其他人的类似。事实上,由于尼安德特人的演化史不同于现代人类,我的这个区域应该与其他人的更为不同。
戴维着手把这些想法用于实际分析。他把人类参考基因组中的欧洲人部分分成片段,然后把这些片段与尼安德特人基因组和其他欧洲人基因组(即克雷格·文特尔的基因组)进行比较,找出差异数目。他发现,总的来说,如果人类参考基因组的欧洲片段与尼安德特人的越接近,那么它们与克雷格的基因组也就越接近。这表明,这些片段中积累的突变,决定了人类参考基因组的欧洲片段与尼安德特人和克雷格基因组的差异。但他发现,在与尼安德特人非常相似的欧洲人片段中,这种关系逆转了,即这些欧洲人基因片段与尼安德特人的越相近,它们与克雷格基因组的差异就越大。虽然根据其他分析,我已相信有基因流发生,但当戴维于 2009 年 12 月访问我们实验室并展示这些结果时,我才确信能够说服世人:尼安德特人的 DNA 片段还留存在当今人类之中。不管怎样研究这些数据,我们都得出了同样的结果。
现在我们可以把注意力转向研究现代人类与尼安德特人以何种方式、在什么时候和什么地方发生密切互动。第一个问题是,基因流的方向:是现代人类贡献 DNA 给尼安德特人,还是尼安德特人贡献 DNA 给现代人类,或两者兼而有之?虽然有人可能认为,当两个人类群体相遇时,基因在两个方向上都是平行流动的,可现实生活中很少有这样的例子,往往是一方占据着社会主导地位。其中一种常见模式:占主导地位的雄性和占非主导地位的雌性交配并产生后代,孩子仍留在母亲所在的非主导群体之中。因此,基因流会从占社会主导地位的群体流向占非主导地位的群体。明显的例子就是美国南部的白人奴隶主,以及非洲和印度的英国殖民主义者。
我们倾向于认为,现代人类比尼安德特人强大,因为尼安德特人最终消失了。但我们的数据却表明,基因流是从尼安德特人流向现代人类的。例如,戴维的最新结果表明,欧洲人与尼安德特人非常相似的 DNA 区域,与其他欧洲人中的那些区域非常不一样。这意味着,在进入现今欧洲人基因库之前的某个时候,这些区域和其他欧洲人的那些区域已经开始分别积累差异。现在假设这种情况同样发生在尼安德特人中。如果贡献是往另一个方向(从现代人类流向尼安德特人),那么这些区域将只是欧洲人基因组的一般区域,差异的数量也应该是其他欧洲人基因组的平均值。基于以上这些分析和其他方面的原因,我们认为所有(或几乎所有)的基因流都是从尼安德特人流入现代人类的。
这并不意味着尼安德特人和现代人类的孩子一定从未由尼安德特人养育。瑞士群体遗传学家洛朗·埃克斯科菲耶一直以来对我们研究组的数据很感兴趣。2008 年,他发表了一篇关于基因流动的论文,文中发生基因流动的双方,其中一方扩张,另一方没有扩张甚至还萎缩了。在这种情况下,种群之间交换得到的基因变异更可能保存在不断扩张的种群中,而非在不断萎缩的种群中。如果基因交换发生在种群发展的“前哨”,而且这个种群正呈现扩张之势,那么交换得来的变异在种群中出现的频率可能会非常高。埃克斯科菲耶曾生动恰当地将这种现象称为“等位基因冲浪”:等位基因进入汹涌的种群扩张浪潮中,使其出现频率激增。这意味着杂交可能在两个方向上均发生过,但我们没有在尼安德特人中发现这一现象,因为现代人类和尼安德特人相遇之后,后者的人口规模已经缩减。
我们之所以无法检测到从现代人类到尼安德特人的基因流,另一个更为现实的原因:凡迪亚洞穴中的尼安德特人生活在 3.8 万年前,那时还没有发生杂交。也许我们永远不会知道尼安德特人与现代人类交配的细节,但这并不困扰我。对我来说,晚更新世的“谁与谁发生性关系”是次要问题。重要的是,尼安德特人真的对当今人类有基因贡献,这可是关乎当今人类遗传起源的问题。
确认了戴维和尼克的研究结果之后,我们开始探讨:非洲以外的人,其基因组有多少来自尼安德特人。这个结果无法通过 SNP 匹配比值直接估计,因为尼安德特人和非洲以外的人的匹配数量取决于许多其他的可变因素:其中一个是尼安德特人和现代人类的共同祖先生活在什么年代,另一个是他们发生杂交的时间,第三个则是尼安德特人曾经的群体规模。蒙蒂·斯拉特金通过模拟尼安德特人和现代人类的群体历史,估计了尼安德特人 DNA在现今人类中的比例。其研究结果表明,欧洲人或亚洲人继承了约 1%~4% 的尼安德特人DNA。戴维和尼克做了不同的分析,他们探索的本质问题是:欧洲人和亚洲人距离百分百的尼安德特人有多远——答案是 1.3%~2.7%。因此,我们的结论是:非洲以外的人的DNA,其中有不到 5% 来自尼安德特人。这个比例虽小,但清晰可辨。
这一轮工作的最后一个问题:除了遗传给欧洲人,尼安德特人又是如何把 DNA 遗传给了中国人和巴布亚人。据我们所知,尼安德特人从没到过中国,也一定没有去过巴布亚新几内亚,所以我们推断,尼安德特人一定是在更偏西边的某个地方与中国人和巴布亚人的祖先相遇。
在每周的电话会议期间,我们围坐在我莱比锡办公室的扬声电话旁。我先把中东的想法压住不表,好让联盟成员用敏锐的头脑去探索所有可能性。蒙蒂想出了一个复杂场景来解释我们看到的变异模式。首先,他认为尼安德特人的祖先原来生活在非洲的某个角落,后来离开了非洲,在大约 40 万至 30 万年前,在欧亚大陆西部演变成尼安德特人。接着,在 20 万年后或更晚的时候,在尼安德特人祖先在非洲起源的地方,现代人类祖先出现了。如果非洲的各种群在这段时间里依然保持分开,那么从尼安德特人的祖先离开、直到现代人类的祖先开始蔓延扩大,等位基因频率的差别就一直存在;然后,现代人类出现之后,不仅走出了非洲,还穿越了非洲,并且与古老的非洲人杂交,获得了早期人类的变异。那么,结果正如我们所看到的那样,相较于非洲人,尼安德特人与非洲以外的人更为相似。
虽然这种情况在理论上是可能的,但前提是,非洲的种群,数万年来保持稳定分开。正如蒙蒂自己指出的,这似乎不可能,因为人类很容易到处移动。更严峻的问题是过往的复杂程度。为了重建过去,最好考虑最简单的模式,即便其他许多更复杂的情况也有可能发生。采用偏好最简单解释的原则,即所谓的简约性(Parsimony)原则。例如,有人假设尼安德特人和现代人类的祖先起源于亚洲,然后现代人类的祖先去了非洲,没有在欧亚大陆留下后裔,随后再次扩张,并取代了尼安德特人。这个假设的确与所有的观察资料吻合,但该假设涉及许多的种群流动和种群灭绝。相对来说,尼安德特人起源于非洲的假说就简单得多。因此相较于非洲起源假说,亚洲起源场景就没那么简约,因此不是当前最好的解释。即便非洲亚结构场景为我们的数据提供了可能的解释,但它依旧不成立,因为有一个更简单、更明显的解释。事实上,这个解释太明显了,我们几个人都不约而同地想道:那就是中东场景(Eastern scenario)。
来源:(瑞典)斯万特·帕博著. 尼安德特人[M]. 杭州:浙江教育出版社, 2018.12.
如何管理大学教师职业行为?明朝一不文明做法,现代老师恐难接受
古代对大学教师品德如何管理?本文作者倪方六“师德”问题,近来备受社会关注。其实,“师德”问题是个老话题,自古就是社会所关心的。外国人会说“教师是人类的灵魂工程师”,中国古人则说老师“为人师表”。中外对教师的评价都很高,自然“师德”就显得十分重要,要求远高于其他职业人士。我要新鲜事2023-05-27 03:46:420000马王堆3号汉墓的这件文物,证明了墓主人身份,还推翻了历史记载
在马王堆1号汉墓的光环下,很多人甚至不知道马王堆3号汉墓。这次的发现大概能让一些人注意到马王堆3号墓。3号墓出土文物也是很丰富的,有千余件,主要有帛画、帛书、简牍、兵器、乐器、漆器、木俑、丝织品等。以下为墓中所出的部分重要文物帛画:T形帛画、车马仪仗图、车马游乐图、导引图帛书:《老子》甲本、乙本、《战国策》、《周易》、《五星占》、《相马经》、《医经方》、《地图》3幅等。我要新鲜事2023-09-09 20:35:330000似奥克龙:北美双足似鸟龙(仅出土盆骨和后肢/疑名)
似奥克龙是一种似鸟龙科恐龙,诞生于8400万年前到6500万年前的白垩纪末期,目前还属于疑名状态,主要使用后两足行走,以肉食为主食,最早的一批化石师在美国出土的,当时仅仅只发现了一块骨盆和一块后肢骨骼。似奥克龙的化石发现我要新鲜事2023-05-10 05:43:440000关于汉代博局纹镜研究的几点思考
汉代是中国古代铜镜发展的繁荣鼎盛时期,在社会生产和生活领域,青铜容器重要性渐次失落,惟铜镜仍为日常不可缺少的必需品而成为当时青铜铸制的重点,能工巧匠集心智与技艺于此,创造出足以夸耀于世的实用工艺品,其中汉代铜镜的研究又至关重要。我要新鲜事2023-05-06 14:21:380000史景迁著作介绍,别样眼光看中国
我要新鲜事2023-05-31 21:02:040000